Wnt訊息傳遞路徑

Wnt訊息傳遞路徑 (Wnt signaling pathway)
國立臺灣大學生命科學系碩士班01級岳威廷
Wnt 蛋白質最早被發現在果蠅的突變品系,這群果蠅的表現型為缺翅型,也因此把此種品系命名為 wingless,而此缺失的蛋白質就被命名為 wingless,但後來經研究發現此基因其實和小鼠的原致癌基因(proto oncogene) int 為同源基因,因此就將這同一群基因命名為 Wnt superfamily。
Wnt 訊息傳遞路徑在動物的分子演化過程中屬於高度保守,目前科學家總共發現 $$13$$ 種不同種類的 Wnt 蛋白質,每種 Wnt 在生物體扮演的角色都不同,而不同種類的 Wnt 蛋白質也會開啟不同的 Wnt 訊息傳遞路徑。
根據路徑開啟時 $$\beta$$-catenin參與的有無,Wnt 訊息傳遞路徑可被分類為典型 Wnt 傳遞路徑(canonical Wnt pathway)及非典型 Wnt 訊息傳遞路徑(non-canonical Wnt pathway),而非典型傳遞路徑又可再被細分為細胞平面極性路徑(cell-planer polarity pathway)以及鈣離子路徑($$\mathrm{Ca^{2+}}$$ pathway)。
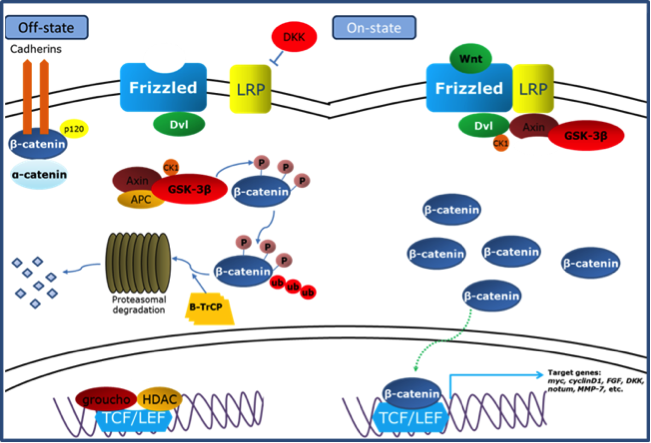
典型 Wnt 訊息傳遞路徑示意圖。(繪圖者:岳威廷。出處:碩士論文)
典型 Wnt 訊息傳遞路徑示意圖說明:Wnt 傳遞路徑關閉:$$\beta$$-catenin 會被 destruction complex(GSK-3$$\beta$$)磷酸化進而被分解,使 $$\beta$$-catenin在細胞質中處於低濃度狀態,讓典型 Wnt 傳遞路徑處於關閉狀態。Wnt 傳遞路徑開啟:當 Wnt 蛋白接上 frizzled 後會活化 dishevelled,dishevelled 會抑制 GSK-3$$\beta$$ 的作用使 $$\beta$$-catenin 能在細胞質累積,一旦 $$\beta$$-catenin 累積足夠濃度便會進入細胞核和 TCF/LEF 作用開啟下游的基因。
以典型 Wnt 傳遞路徑為例(如圖例),在 Wnt 傳遞路徑尚未開啟時,$$\beta$$-catenin即會被細胞常態表現在細胞質中。當細胞質中的 axin、APC 及 GSK-3$$\beta$$ 等蛋白質結合形成 destruction complex 時,GSK-3$$\beta$$ 會對 $$\beta$$-catenin 進行磷酸化,使 $$\beta$$-catenin 先接上泛素(ubiquitin),最後被蛋白酶分解,此時細胞質中的 $$\beta$$-catenin 處於低濃度的狀態,Wnt signaling 因而無法被開啟。
但是當 Wnt ligand 接上其受體(receptor)frizzled 後,會和 LRP 一起活化下游的蛋白質 dishevelled,dishevelled 被活化後會促進 destruction complex 的分解,使 GSK-3$$\beta$$ 無法對 $$\beta$$-catenin 進行磷酸化,這時 $$\beta$$-catenin 便能在細胞質中累積,當濃度到達一定程度後,$$\beta$$-catenin便會進入細胞核,和轉錄因子「T 細胞因子/淋巴增強因子(TCF/LEF)」作用開啟下游的基因(例如:Dickkopf、Myc、FGF…),此步驟便是典型 Wnt 傳遞路徑開啟的過程。
在傳遞過程中無論屬於何類傳遞路徑,Wnt、frizzled 及 dshevelled 為所有 Wnt 傳遞路徑都共同需要參與的蛋白質。
根據目前研究,能調控 Wnt 傳遞路徑的方式有很多,例如 dickkopf 蛋白質也能和 Frizzled 接合,跟 Wnt 蛋白質競爭受體達到拮抗的作用。而 Hedgehog 訊息傳遞路徑則被發現是 Wnt 傳遞路徑的上游,會促進 Wnt 訊息傳遞路徑的開啟。此外,BMP 訊息傳遞路徑也被發現會透過調控 GSK-3$$\beta$$ 和 Wnt 傳遞路徑達到橫向連接對話(cross talk)的效果。
Wnt 傳遞路徑最早被發現和細胞癌化有關,接著被發現和胚胎發育、體軸建立、細胞遷移、細胞分裂、細胞分化及幹細胞全能性等皆有關。
在細胞株的實驗中,Wnt 傳遞路徑被發現會調控細胞分裂、細胞分化及細胞遷移,因此若 Wnt 訊息傳遞路徑調控異常有可能會導致癌化;在幹細胞的研究中,Wnt 則被指出對幹細胞的細胞分裂及全能性有關。
除了在細胞層次的研究,Wnt 訊息傳遞路徑在動物個體扮演的角色也被廣泛的研究討論,在果蠅的胚胎發育實驗中,Wnt 被發現和背腹(dorsal/ventral body axis)的體軸建立有關。
近來,Wnt 傳遞路徑也被發現和前後體軸的建立(anterior/posterior body axis)有關,在渦蟲再生時若抑制典型 Wnt 傳遞路徑會使渦蟲成為雙頭或多頭但無尾的表現型,而促進典型 Wnt 傳遞路徑開啟則會造成雙尾無頭的表現型,證實 Wnt 訊息傳遞路徑在前後體軸的建立扮演重要的角色。而在小鼠的實驗中,Wnt 傳遞路徑也和腦神經的發育有關。另外在雞和小鼠的實驗中,Wnt 訊息傳遞也被發現和性腺發育有關。
註解
- 原致癌基因(proto-oncogene),屬於原本細胞內舊有的正常基因,但是會因為突變或被病毒攜帶等因素轉變成致癌基因(oncogene)。
- $$\beta$$-catenin 除了參與 Wnt 傳遞路徑外,還會和 cadherin 及 $$\alpha$$-catenin 結合形成黏合接點(adherens junction)複合體,功能為建立細胞間的彼此連結。
參考文獻:
- Andersson, E. R., Saltó, C., Villaescusa, J. C., Cajanek, L., Yang, S., Bryjova, L., . . . Arenas, E. (2013). Wnt5a cooperates with canonical Wnts to generate midbrain dopaminergic neurons in vivo and in stem cells. Proceedings of the National Academy of Sciences, 110(7), E602–E610. doi: 10.1073/pnas.1208524110
- Lander, A., Kimble, J., Clevers, H., Fuchs, E., Montarras, D., Buckingham, M., . . . Oskarsson, T. (2012). What does the concept of the stem cell niche really mean today? BMC Biology, 10(1), 19.
- Lim, C.-H., Lim, W., Jeong, W., Lee, J.-Y., Bae, S.-M., Kim, J., . . . Song, G. (2013). Avian WNT4 in the Female Reproductive Tracts: Potential Role of Oviduct Development and Ovarian Carcinogenesis. PLoS ONE, 8(7), e65935. doi: 10.1371/journal.pone.0065935
- Moore, K. A., & Lemischka, I. R. (2006). Stem Cells and Their Niches. Science, 311(5769), 1880-1885. doi: 10.1126/science.1110542
- Nusse, R. (2012). Wnt Signaling. Cold Spring Harbor Perspectives in Biology, 4(5). doi: 10.1101/cshperspect.a011163
- Strochlic, L., Falk, J., Goillot, E., Sigoillot, S., Bourgeois, F., Delers, P., . . . Legay, C. (2012). Wnt4 Participates in the Formation of Vertebrate Neuromuscular Junction. PLoS ONE, 7(1), e29976. doi: 10.1371/journal.pone.0029976
- 岳威廷。2014年。經典Wnt/β-catenin訊息傳遞路徑相關基因在瓢體蟲(Aeolosoma viride)前端再生的表現。國立臺灣大學生命科學系碩士論文。
 前一篇文章
前一篇文章 下一篇文章
下一篇文章 如何防止自體免疫反應
如何防止自體免疫反應 .jpg) 【探索基礎科學系列講座19】翻轉吧,DNA ∞ 雙螺旋65歲生日趴
【探索基礎科學系列講座19】翻轉吧,DNA ∞ 雙螺旋65歲生日趴  頭皮毛囊與精神疾病診斷
頭皮毛囊與精神疾病診斷  細胞內測量長度的蛋白質
細胞內測量長度的蛋白質  細胞膜的構造
細胞膜的構造 ![[新聞] 發現臺灣目前最古老人類化石](https://highscope.ch.ntu.edu.tw/wordpress/wp-content/uploads/2015/01/pe.png) [新聞] 發現臺灣目前最古老人類化石
[新聞] 發現臺灣目前最古老人類化石  年齡提高卵母細胞染色體異常機率
年齡提高卵母細胞染色體異常機率  伊波拉病毒(Ebola virus)—病毒的防制
伊波拉病毒(Ebola virus)—病毒的防制 

