雌性選擇(Female choice):面天樹蛙雄蛙重疊鳴叫對雌性選擇的影響

雌性選擇(Female choice):面天樹蛙雄蛙重疊鳴叫對雌性選擇的影響
國立清華大學生命科學系學士班2012級辜昱嘉
除了生存競爭的「天擇」,擇偶也是演化動力的來源,雄性間競爭與雌性的交配機會,失敗者並非死亡,只是在子代數目上吃虧。然而往往雌性才具有生殖主導權,雌性從眾多雄性中選擇配偶以繁殖基因優良的子代,或僅依雌性本身的狀況(如:存活力、生殖力)做選擇(Bourne, 1993)。
兩棲類以聽覺作為主要溝通媒介,發出鳴叫聲來傳達訊息。求偶叫聲可透露雄性品質(如:體型、年齡)與位置,雌蛙可依據雄蛙鳴叫聲傳送的訊息來選擇雄蛙進行交配。而當有兩隻以上個體鳴叫時,個體鳴叫的時間點會影響鳴叫是否重疊。在多數自然情況下,雄蛙彼此會錯開鳴叫,如此雄性較容易由鳴叫聲去衡量競爭者的優勢程度(Schwartz, 1987)。
具有聚集展示(lek)行為的蛙類,雄蛙會聚集在一個展示場鳴叫吸引雌蛙靠近,整個生殖過程中雄蛙並不提供除生殖細胞外的任何資源給雌蛙,雌蛙主要靠雄蛙的鳴叫聲來選擇雄性。而具有領域性的蛙類,雄蛙會藉由打鬥佔據較好的生殖位置以吸引雌性,對雌蛙而言,選擇的依據不單只是雄蛙的鳴叫聲,還有雄蛙所掌控的環境資源,如生殖棲位,因此鳴叫聲所代表的雄性優勢程度就不再是雌蛙進行雌性選擇時唯一、且最重要的參考要素。
面天樹蛙(Kurixalus idiootocus)求偶時不會刻意的防衛固定領域(張, 1989),而是採取彼此間達最小空間(nation minimum inter-male space)的策略,雄蛙彼此競爭的僅是鳴叫機會,而不是鳴叫位置本身(Wells, 1977),屬於聚集展示的物種。當一群雄蛙聚集在一起鳴叫時,鳴聲干擾甚大,時常有鳴聲重疊的情形,面天樹蛙雄蛙在展示場內的重疊鳴叫是否會干擾雌性對鳴叫聲傳遞訊息的判別?以下實驗可以探討雄蛙重疊鳴叫是否會對雌性選擇造成影響。

圖一、實驗裝置圖
在實驗區域兩端各放置一個音箱,連接錄音筆,分別以左右聲道播出不同處理的聲音,中央譜架下放置一以棉線連接之塑膠蓋,右上角為實驗場域分割之示意圖。(照片與繪圖:辜昱嘉)
將實驗場域分割為四個象限(見圖一),第I、II象限為有效選擇區,第II、IV象限為無效選擇區,在橫跨第I、III象限的直徑兩端各置一音箱連接音源線至錄音筆,分別輸出左右聲道播放處理音檔。
為模擬夜間環境,僅以前端套有紅色玻璃紙的手電筒作為光源,將面天樹蛙雌蛙置於中央塑膠蓋下,播放5分鐘音檔使其適應後,由遠處拉引棉線使塑膠蓋沿譜架垂直上升,同時持續重複播放音檔,觀察雌蛙動向,當雌蛙離開實驗場域,觸碰到場域邊界由粉筆繪製的線條時即代表實驗結束,記錄雌蛙離開實驗場域邊界之所屬象限,若個體在30分鐘內未離開場域即表示實驗失敗。
在分別以十七隻面天樹蛙雌蛙進行不同音檔處理(剪輯或重疊面天樹蛙雄蛙音訊,包括鳴叫間隔、鳴叫連續、相同鳴叫50%重疊、相異鳴叫50%重疊等,依實驗設計取兩者作次序搭配)實驗後,發現雌蛙偏好選擇重疊的雄性鳴叫(影片連結:http://youtu.be/maef0ldp7dU,影片於00:04後為十倍速快轉,可明顯看出面天樹蛙能對鳴叫聲做出選擇)。
因此,推論影響雌性選擇的主因可能是雄蛙的聚集程度,雌蛙將不同雄蛙的重疊鳴叫視為多個雄性聚集處,故偏好雄蛙重疊鳴叫。鳴叫聲的重疊並不干擾雌蛙做選擇,重疊的鳴叫聲皆是雄性面天樹蛙的鳴叫聲,同種雄性的鳴叫聲對雌性而言可能有放大加成的效果,讓雌蛙認為那個方位具有較多雄性。這是對雌性有利的行為,因為雄性聚集的場所雌性可以較快找到配對的雄性,也有較高的機會找到較優秀的雄性。
然而若是以異種雄性的鳴叫聲進行重疊,可能對雌蛙而言是聽覺上的干擾,進而選擇沒有重疊的方位,然而現階段仍缺乏相關研究驗證此假說。
參考文獻:
- 張耀文(1989)。面天樹蛙生殖行為之研究。國立臺灣大學動物所碩士論文。
- Bourne, Godfrey R. (1993). Proximate costs and benefits of mate acquisition at leks of the frog Ololygonrubra. Animal Behaviour, 45(6), 1051-1059.
- Grafe, T. U. (1997). Costs and benefits of mate choice in the lek-breeding reed frog Hyperolius marmoratus. Animal Behaviour, 53(5), 1103-1117.
- Hutchinson, J. M. C. (2005). Is more choice always desirable? Evidence and arguments from leks, food selection, and environmental enrichment. Biological Reviews, 80: 73-92. doi: 10.1017/S1464793104006554
- Kirkpatrick, M. & Ryan, Michael J. (1999). The evolution of mating preferences and the paradox of the lek. Nature 350, 33-38.
- Kuramoto, M., & Wang, C. S. (1987). A new rhacophorid tree frog from Taiwan, with comparisons to Chirixalus eiffingeri (Anura, Rhacophoridae). Copeia, 1987, 931-942.
- Robertson, J. G. (1986). Female choice, male strategies and the role of vocalizations in the Australian frog Uperoleiarugosa. Animal Behaviour, 34(3), 773-784.
- Schwartz, J. J. (1987). The function of call alternation in anuran amphibians: a test of three hypotheses. Evolution, 41(3), 461-471.
- Weale, R. A. (1956). Observations on the direct effect of light on the irides of Ranatemporaria and Xenopuslaevis. The Journal of physiology, 132(2), 257-266.
- Wells, K. D. (1977). The social behaviour of anuran amphibians. Animal Behaviour, 25, 666-693.
- Wells, K. D., & Schwartz, J. J. (2006). The behavioral ecology of anuran communication. In Hearing and sound communication in amphibians (pp. 44-86). Springer New York.
 前一篇文章
前一篇文章 下一篇文章
下一篇文章 【2013諾貝爾獎特別報導】生醫獎:破解細胞傳輸系統之謎
【2013諾貝爾獎特別報導】生醫獎:破解細胞傳輸系統之謎  白蟻透過腸道微生物高效率獲得能量
白蟻透過腸道微生物高效率獲得能量  細胞老化誘發腫瘤化
細胞老化誘發腫瘤化  戀愛活化大腦多巴胺神經
戀愛活化大腦多巴胺神經  【2015諾貝爾生醫獎特別報導】阿弗麥克素及青蒿素-對抗寄生蟲疾病的革命性治療法(一)
【2015諾貝爾生醫獎特別報導】阿弗麥克素及青蒿素-對抗寄生蟲疾病的革命性治療法(一) 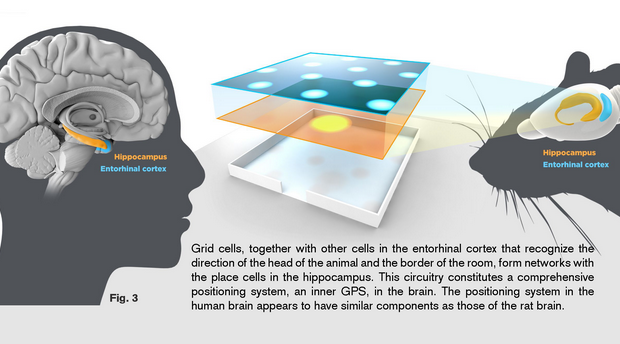 【2014諾貝爾生醫獎】發現大腦裡空間記憶的構築細胞
【2014諾貝爾生醫獎】發現大腦裡空間記憶的構築細胞 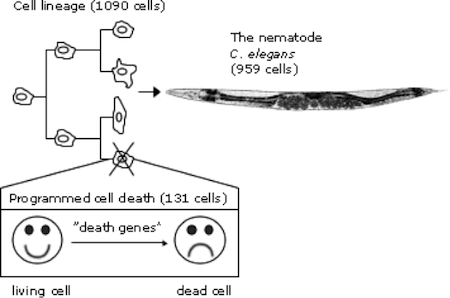 細胞凋亡 (Apoptosis)
細胞凋亡 (Apoptosis)  治療乳癌末期新藥 FDA批准上市
治療乳癌末期新藥 FDA批准上市 

