潛水反射

潛水反射 (Diving reflex)
國立臺灣大學生態學與演化生物學碩士 陳雪溱
潛水反射出現於所有目前已測試過的海洋與陸生脊椎動物,也包含人類。目的為在潛水的過程中降低四肢肌肉與腹部器官的氧氣消耗,將氧氣保留給大腦、心臟與肺,延緩可能的大腦損傷直至呼吸恢復。潛水反射在需要長時間憋氣以進行捕食或活動的海洋哺乳動物身上尤其明顯,例如海豚、鯨魚與海豹等。
潛水反射在生理上主要有四項:
- 心跳減慢 (Bradycardia)
在臉部浸入比體溫低的水中時,屬於副交感神經系統 (Parasympathetic nervous system) 的三叉神經 (Trigeminal nerve) 中的眼神經 (Ophthalmic branch) 與上頷神經 (Maxillary branch) 的冷受器受到刺激,進一步激發迷走神經運動神經元 (Vagal motoneurons) 使心跳速率降低 (Shattock and Tipton, 2012)。在時常進行自由潛水的人類身上大約會降低至平時的 50%,而生活在海洋的動物此反應更加明顯。加州一項實驗發現,7 頭小海象 (Odobenus rosmarus) 在陸地上的平均心跳為每分鍾 107 次,當牠們在海水中下潛時,心跳下降到平均每分鍾 39 次,下降了 64%。而皇帝企鵝 (Aptenodytes forsteri) 在潛水過程中的心跳速率甚至會降到每分鐘 3 次。
- 末梢血管收縮 (Peripheral vasoconstriction)
在心跳速率降低的同時,動脈血壓會下降,引起交感神經系統 (Sympathetic nervous system) 作用,使末梢血管收縮,如皮膚、四肢肌肉、內臟器官的血流量將減少。除了有維持核心溫度體溫的效果外,末梢血管收縮會將血液保持在身體核心,將氧氣優先提供給大腦、心臟與肺等重要器官 (Andersson et al., 2002)。
- 脾臟收縮 (Spleen contraction)
此反應在水生哺乳類動物首先被發現,經由交感神經系統作用引發。海洋哺乳動物擁有較大比例的脾臟,在下潛時,脾臟會收縮並將紅血球釋放進血管通路中,使得血液中攜帶的氧氣量增加。威德爾海豹 (Leptonychotes weddellii) 在下潛的 10-12 分鐘時,其紅血球容積比 (Hematocrit) 與血紅素濃度 (Hemoglobin concentration) 分別上升了 48% 和 44% (Qvist et al., 1986)。Schagatay 等人 (2001) 進行的研究中, 20 位受試者皆未受過專業的憋氣訓練,其中有半數曾經接受過脾臟移除手術,受試者將臉浸入 10°C 冷水中閉氣 5 次,結果顯示有脾臟的受試者其紅血球容積上升了 6.4%、血紅素濃度上升了 3.3%,顯著高於脾臟移除的受試者。雖然人類脾臟對氧氣的供應比例相較海洋哺乳動物相差甚遠,但是此研究證實了人類的潛水反應也包含了脾臟收縮。
- 血液轉移 (Blood shift)
另外一項因潛水深度造成的生理變化為胸腔的血液轉移 (Blood shift),在憋氣下潛的活動中(如海女、自由潛水者),肺泡內的空氣體積被壓縮,且隨下潛深度加大空氣體積持續下降,此時肺部血管血壓上升,使肺泡周圍的微血管充血膨脹(圖一),以維持肺的形狀使之不因其體積下降與肋膜腔 (Intrapleural) 拉扯、分離使肺泡破裂而受傷。哺乳動物的胸腔是由橫隔膜、胸骨與肋骨所構成,肺與胸腔之間的空間稱作肋膜腔,正常情況下肋膜腔內沒有空氣且為負壓狀態,使得胸腔的擴張可以帶動肺膨脹吸氣。胸腔體積的變動程度隨物種不同差異甚大,例如下潛的最大深度可達 3000 公尺的抹香鯨 (Physeter macrocephalus),其大多數的肋骨與胸骨分離,使得胸腔有較大的活動空間,在深海中受到強大水壓會隨之縮小。
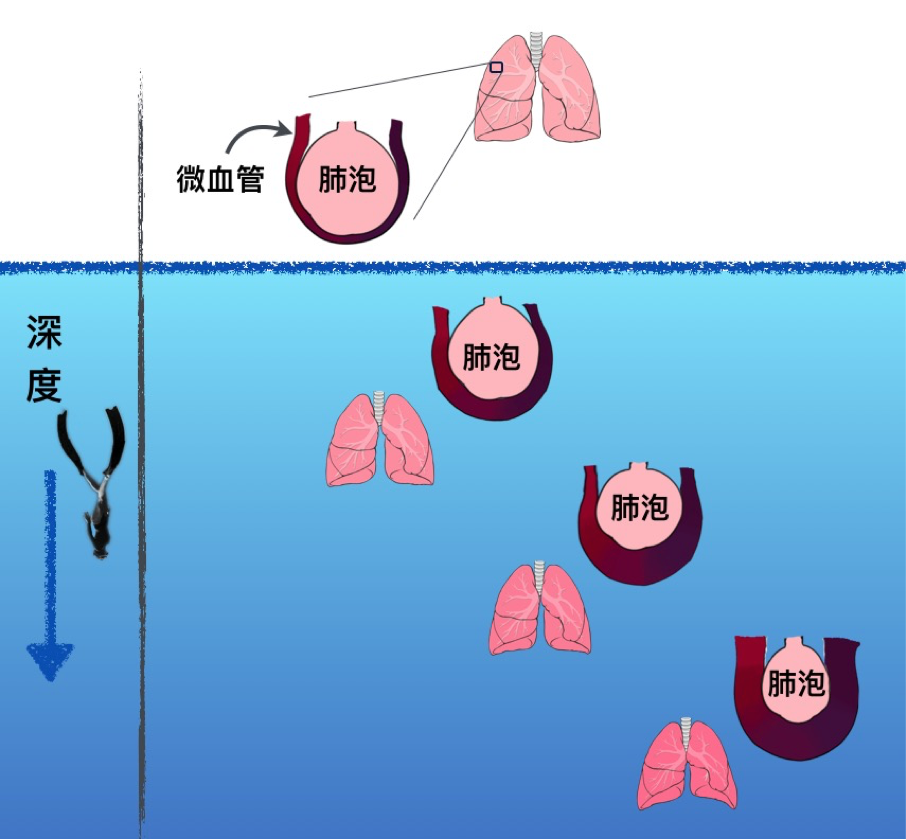
圖一、血液轉移 (blood shift),肺泡周圍的微血管隨著下潛深度而逐漸膨脹充血。因水壓壓縮肺內空氣使肺被壓縮,充血的微血管有支持肺形狀的功能,但血液仍在血管內,並沒有進入肺泡中。(本文作者陳雪溱繪)
然而相較之下,人類的胸腔活動程度較小,在憋氣下潛水深超過胸腔能被壓縮的極限後,肺內的空氣體積若持續下降,肋膜腔被拉扯將會造成肺泡破裂,因此在 1960 年以前,科學家相信人類不可能以一口氣潛水超過 50 公尺,不是因為憋氣時間不足,而是肺內空氣體積受到水壓擠壓下降造成的肺部損傷。但爾後不斷有自由潛水者打破此紀錄並毫髮無傷地回到水面,研究者透過自由潛水者在不同水深的肺容積,發現人類胸腔經由血液轉移填補的血液流量可達約 1 公升,由於液體受到壓力時體積的變化很小,這些轉移進胸腔血管的額外血液便能達到維持肺形狀的作用,在浮出水面的過程中,血液便會透過血管隨水壓減小而離開胸腔 (Craig et al., 1968;Schaefer et al., 1968)。但是在極端深度與低氧的狀況下,因為微血管過度膨脹,曾有血液中的血漿滲出至肺泡中導致肺水腫 (Pulmonary edema) 或出血 (Hemorrhage) 的案例 (Linér and Andersson, 2008)。水肺潛水者不會有此反應,因為氣瓶中的高壓氣體可以供給胸腔所需要的空氣壓力以維持其體積。
另外,中耳腔室是另一個需要平衡的部位,人類在進行大深度潛水時主要以空氣進行壓力平衡,但部分海洋哺乳動物是以血液轉移的機制來進行中耳腔室的平衡 (Costa, 2007)。
參考文獻
- Andersson, J. P., Linér, M. H., Rünow, E., & Schagatay, E. K. (2002). Diving response and arterial oxygen saturation during apnea and exercise in breath-hold divers. Journal of Applied Physiology, 93(3), 882-886.
- Craig Jr, A. B. (1968). Depth limits of breath hold diving (an example of Fennology). Respiration Physiology, 5: 14-22.
- Costa, D. P. (2007). Diving Physiology of Marine Vertebrates. University of California Museum of Paleontology. Retrieved from http://www.ucmp.berkeley.edu/about/shortcourses/costa_divingphysiology2007.pdf
- Linér, M. H. and Andersson, J. P. (2008). Pulmonary edema after competitive breath-hold diving. Journal of Applied Physiology, 104: 986–990.
- Qvist, J. E. S. P. E. R., Hill, R. D., Schneider, R. C., Falke, K. J., Liggins, G. C., Guppy, M. I. C. H. A. E. L., … & Zapol, W. M. (1986). Hemoglobin concentrations and blood gas tensions of free-diving Weddell seals. Journal of Applied Physiology, 61(4), 1560-1569.
- Schagatay, E., Andersson, J. P., Hallén, M., & Pålsson, B. (2001). Selected contribution: role of spleen emptying in prolonging apneas in humans. Journal of Applied Physiology, 90(4), 1623-1629.
- Schaefer, K. E., Allison, R. D., Dougherty Jr, J. H., Carey, C. R., & Walker, R. (1968). Pulmonary And Circulatory Adjustment Determining The Limits Of Depths In Breathhold Diving. Science 162: 1020-1023.
- Shattock, M. J., & Tipton, M. J. (2012). ‘Autonomic conflict’: a different way to die during cold water immersion?. The Journal of physiology, 590(14), 3219-3230.
 前一篇文章
前一篇文章 下一篇文章
下一篇文章 細胞內測量長度的蛋白質
細胞內測量長度的蛋白質  憂鬱症治療新契機
憂鬱症治療新契機  細胞膜的構造
細胞膜的構造  【2016年諾貝爾生理醫學獎特別報導】發現自噬作用機制
【2016年諾貝爾生理醫學獎特別報導】發現自噬作用機制 ![[講座] 尼安德塔人是什麼樣的人?](https://highscope.ch.ntu.edu.tw/wordpress/wp-content/uploads/2015/02/尼安德塔1.png) [講座] 尼安德塔人是什麼樣的人?
[講座] 尼安德塔人是什麼樣的人?  伊波拉病毒(Ebola virus)—病毒的防制
伊波拉病毒(Ebola virus)—病毒的防制  負責運動學習的蛋白質
負責運動學習的蛋白質  國際生物多樣性日專題 – 何謂生物多樣性?
國際生物多樣性日專題 – 何謂生物多樣性? 

