致癌基因 ─中
致癌基因 (Oncogene) ─中
國立臺灣師範大學生命科學系碩士生黃培綺
原致癌基因參與我們正常細胞生命週期的階段,包括在細胞生命中的「細胞生長」、「細胞分裂(cell division)」和「細胞分化(cellular differentiation)」等這三個階段。在某些特定情況下,像是長期接觸到致癌物質(carcinogen),例如致突變劑(mutagen)、輻射或放射性同位素(radionuclide)等,或是細胞被病毒(virus)感染,這些外來因素影響,會使正常細胞的DNA受到損害或基因表現量異常,若其結果導致原致癌基因發生突變(mutation)或基因大量表現的話,導致「致癌基因」發生,這樣就會進一步影響正常細胞的生命週期。
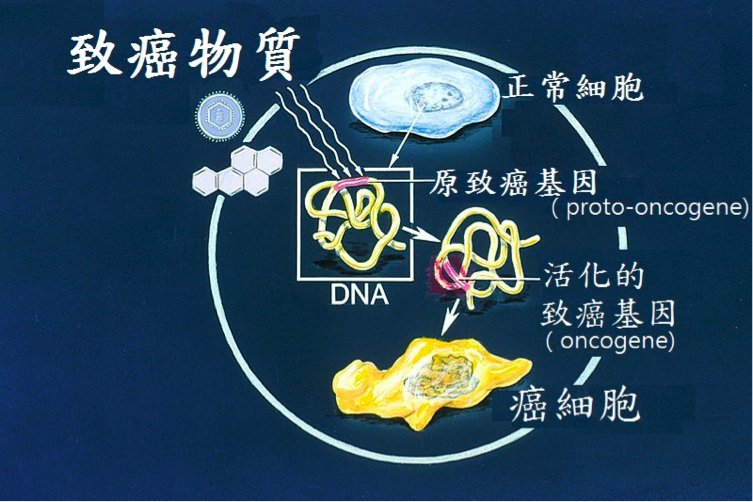
圖三、當細胞接受刺激使致癌基因被活化後,正常細胞轉變為癌細胞的示意圖。(翻譯自File:Oncogenes illustration.jpg – Wikimedia Commons. http://commons.wikimedia.org/wiki/File:Oncogenes_illustration.jpg)。
正常的細胞除了幹細胞外,終其一生,其細胞生長、分裂的次數是有限制的,而且在細胞生命週期的最後,細胞會老化,最後死亡。